Kamoun Ophtalmology
Anatomie de l'oeil
L'oeil, qui est l'organe de la vision et est de faible volume (6.5 cm3), pèse 7 grammes, et a la forme d'une sphère d'environ 24 mm de diamètre, complétée vers l'avant par une autre demi-sphère de 8 mm de rayon, la cornée.
La paroi du globe oculaire est formée de
3 tuniques :
- La tunique fibreuse, externe, se compose de la sclérotique opaque en arrière et de la cornée transparente en avant. La sclérotique : c'est une membrane rigide qui donne sa forme à l'oeil. Devant, elle devient la cornée. Membrane fibreuse, résistante, elle entoure et protège l'oeil à l'extérieur tout en maintenant sa forme. La sclérotique forme ce que l'on appelle couramment "le blanc de l'oeil". Elle est traversée par un grand nombre de petits canaux (artères, nerfs, veines) et, à l'arrière, par une ouverture où passent les fibres du nerf optique et qui s'appelle la lame criblée. La cornée constitue la lentille principale du système optique oculaire ; pour que ce tissu puisse remplir sa fonction il doit être transparent et tout concourt à assurer cette transparence. La cornée est enchâssée dans l'ouverture antérieure de la sclérotique. La frontière entre cornée et sclérotique, appelée limbe, est une zone semi-transparente qui a la particularité d'adhérer à la conjonctive, fine membrane qui couvre la face interne des paupières et la portion antérieure de la sclérotique. Tandis que la cornée est avasculaire, le limbe est richement innervé et vascularisé.
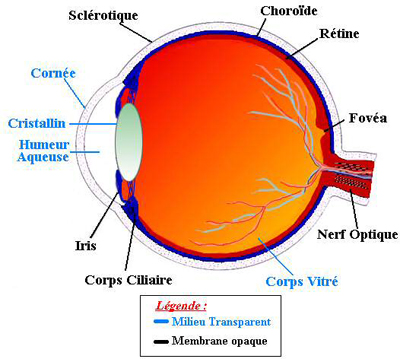
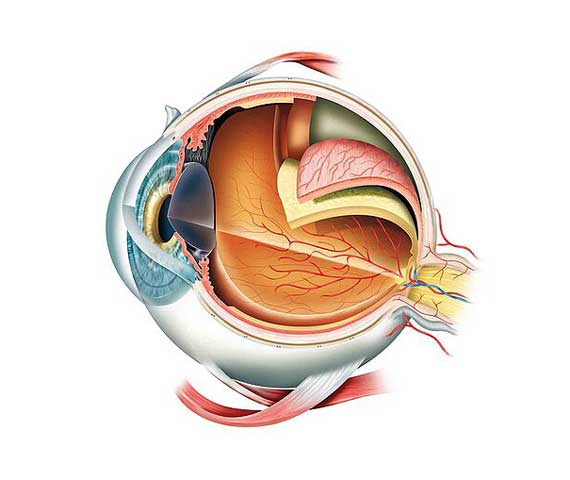
- La tunique uvéale, dite aussi uvée, se compose de trois éléments : l'iris en avant, le corps ciliaire et la choroïde en arrière. L'iris est la portion la plus antérieure de l'uvée; il est de structure pigmentée, donnant sa couleur à l'oeil et percé d'un trou, la pupille. La contraction ou la dilatation réflexes règlent la quantité de lumière pénétrant dans l'oeil par la pupille. Il est situé dans l'humeur aqueuse entre la cornée et le cristallin, séparant ainsi la chambre antérieure de la chambre postérieure de l'oeil. L'iris est innervé par des fibres du système nerveux qui activent les muscles dilatateurs et sphincters responsables de la dilatation et de la constriction pupillaires. Le corps ciliaire est représenté par un épaississement de l'uvée situé en couronne derrière l'iris. C'est un anneau de tissu musculaire qui produit une substance liquide appelée humeur aqueuse ; ses muscles maintiennent le cristallin, et en modifient la forme. La choroïde est une couche vasculaire, tapissant l'intérieur du globe. C'est le tissu nourricier de l'oeil: il apporte l'oxygène et les nutriments dont les cellules ont besoin pour leur métabolisme.
- La tunique nerveuse se compose de la rétine, que nous étudierons précisément plus tard. C'est elle qui traduit les rayons lumineux en influx nerveux. Ces tuniques enferment des milieux transparents, indispensables à la vision. Ces milieux transparents sont au nombre de trois et forment une lentille convergente.
- L'humeur aqueuse : c'est un liquide transparent qui remplit l'espace entre la cornée et le cristallin, liquide continuellement renouvelé et qui, avec le corps vitré, maintient la pression oculaire. S'il y a trop de pression oculaire, il y a une mauvaise irrigation sanguine de la papille, le lieu d'émergence du nerf optique.
- Le cristallin : c'est la lentille de l'oeil qui permet la mise au point par sa propriété essentielle qu'est sa plasticité qui lui permet de focaliser la lumière sur la rétine en modifiant ses courbures lors de l'accommodation. De forme biconvexe, flexible et transparent, il est situé à l'intérieur du globe oculaire. Sa partie antérieure est en contact avec l'humeur aqueuse et sa partie postérieure avec le corps vitré. Il est maintenu en place par une série de fibres amarrées au corps ciliaire, la zonule de Zinn.
- Le corps vitré : c'est une masse gélatineuse et transparente, contenant 99% d'eau et représentant 60% du volume oculaire. Le corps vitré maintient la rétine contre les parois de l'oeil. Nous venons donc de voir que l'oeil est constitué de trois membranes: la sclérotique, la choroïde et la rétine, et de trois milieux transparents: l'humeur aqueuse, le cristallin et le corps vitré. Les membranes, hétérogènes et opaques (sauf pour la cornée), sont à la périphérie de l'oeil. En revanche, les milieux transparents, homogènes, sont au centre de l'oeil, garantissant l'acheminement des rayons lumineux à l'écran rétinien.
La rétine
- Tapissant le fond de l'oeil, la rétine visuelle ou nerveuse est le lieu de traduction du message lumineux venant de l'extérieur en signaux nerveux envoyés au cerveau. Il s'agit d'un tissu neuronal très fin, qui fait partie du Système Nerveux Central, de 0,1 à 0,5 mm d'épaisseur, organisée en dix couches de cellules. Celles-ci comprennent l'épithélium pigmentaire, la couche des photorécepteurs, la membrane limitante, la couche granuleuse et la couche plexiforme externes et internes, la couche des ganglionnaires, la couche des fibres optiques et finalement la membrane limitante interne. Pour simplifier, nous parlerons ici uniquement de trois couches qui nous intéressent: la couche des cellules photosensibles comprenant la couche des photorécepteurs et la couche granuleuse externe, la couche granuleuse interne et la couche des ganglionnaires. Ces trois couches représentent les cellules nerveuses de la rétine, reliées entre elles et ayant chacune une fonction précise.
- La plus profonde, par rapport à l'arrivée de la lumière est, paradoxalement, la couche des cellules photosensibles, aussi appelées photorécepteurs ou cellules sensorielles. La rétine est en effet "inversée", car la lumière doit traverser la rétine avant de pouvoir atteindre les photorécepteurs, sensibles à la lumière. Cette couche comporte environ 130 millions de cellules photosensibles différentes, portant des noms reflétant leur forme:
- Les bâtonnets, qui constituent environ 95% de ces cellules, soit au nombre de 120 millions, sont responsables de la vision nocturne, et ne sont sensibles qu'à la différence entre obscurité et lumière. Par contre, ils ont la plus grande sensibilité, et sont par cela adaptés à de faibles quantités de lumière.
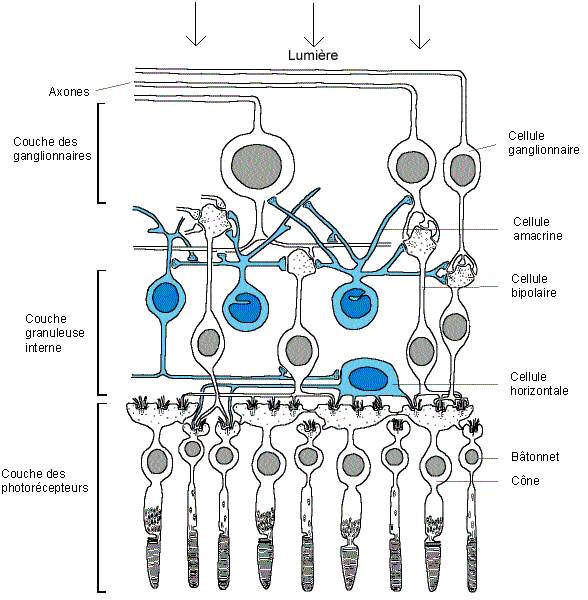
- D'autre part, les cônes, cellules sensorielles plus grandes, forment les 5% restants des photorécepteurs. Les cônes sont responsables de la vision diurne (de jour), et font la différence entre les couleurs. Les cônes sont présents en majorité au niveau de la "tache jaune", dans la région centrale de la rétine. Au milieu de celle-ci se trouve la "fovéa", une légère dépression où sont présents uniquement des cônes, très serrés, et où les autres couches sont rejetées à la périphérie, pour laisser pénétrer la lumière plus facilement. La vision est à ce niveau plus précise, plus détaillée et plus sensible aux mouvements que sur le reste de la rétine. C'est d'ici que provient la plupart de l'information visuelle arrivant au cerveau.
- La couche suivante, dénommée couche granuleuse interne, comporte une variété de neurones rétiniens, qui peuvent être classés en trois catégories:
- Les cellules bipolaires, tenant leur nom du fait qu'elles soient articulées entre les photorécepteurs et les cellules ganglionnaires de la 3e couche (les deux "pôles"), constituent la voie "directe" de transmission du message nerveux. Il existe plusieurs groupes de cellules bipolaires: les bipolaires de bâtonnets, reliant plusieurs bâtonnets à une cellule ganglionnaire, et les bipolaires de cônes, reliant un ou plusieurs cônes à une cellule ganglionnaire. Ces derniers se subdivisent encore en deux groupes, les bipolaires "invaginées" et les bipolaires "superficielles", formant deux voies distinctes de transmission de l'information. Les deux autres groupes de neurones de cette deuxième couche servent à moduler latéralement la transmission d'information, formant un "système de contrôle" qui permet au système rétinien de tenir compte des évènements issus du voisinage, pour mieux s'adapter, par exemple, aux contrastes ou aux bords des objets, et aussi au mélange des couleurs.
- Les cellules horizontales, dont le nom décrit bien le placement, en contact avec les synapses (région de contact entre deux neurones, et lieu de transfert d'informations entre celles-ci) entre les récepteurs et les cellules bipolaires.
- Les cellules horizontales, dont le nom décrit bien le placement, en contact avec les synapses (région de contact entre deux neurones, et lieu de transfert d'informations entre celles-ci) entre les récepteurs et les cellules bipolaires.
- Les cellules amacrines, en contact avec les synapses entre cellules bipolaires et ganglionnaires. La dernière couche nerveuse de la rétine, "l'étage de sortie" vers le cerveau, est composée d'environ 1 million de cellules ganglionnaires, soit à peu près 130 fois moins nombreux que les cellules photosensibles. Ces neurones sont reliés d'une part aux cellules bipolaires, et ont d'autre part des fibres nerveuses, ou axones, qui se rejoignent au niveau de la papille pour former le nerf optique, relié au cerveau. La papille est dépourvue de photorécepteurs, et forme donc une "tache aveugle". Ceci n'entraîne pourtant pas de problème visuel, grâce à une compensation de la part du cerveau. Grâce à cette organisation en couches, l'information est donc véhiculée vers le cerveau en plusieurs étapes, ce qui permet d'effectuer un traitement des signaux au niveau rétinien; nous verrons plus tard comment.
Les cônes
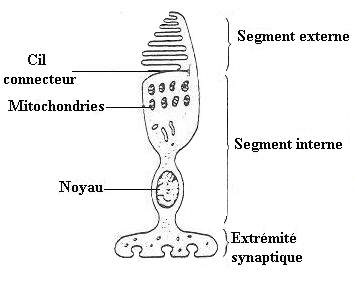
- Au nombre de 6 millions, engouffrés dans la rétine, les cônes jouent un rôle fondamental dans la perception des couleurs. Le nom de "cône" leur est attribué car ces cellules en ont la forme. Un cône est constitué de deux parties distinctes: un segment externe et un segment interne, ces deux parties étant reliées par un cil connecteur.
- Le segment interne contient le noyau et les organites (les mitochondries, l'appareil de Golgi, etc.) indispensables au fonctionnement de toute cellule.
- Dans sa partie la plus distale, son pied, de taille relativement grande, le segment interne possède divers types de synapses: des synapses électriques, où le transfert du message nerveux est comparable à une simple conduction électrique, assurent des relations entre photorécepteurs voisins (cônes et bâtonnets), et des synapses chimiques qui permettent la transmission du message nerveux à l'aide d'un neuromédiateur; il s'agit dans le cas présent d'un acide aminé: le glutamate. Celui-ci est contenu dans le bouton synaptique (renflement du prolongement pré-synaptique) et sera expulsé de la membrane pré-synaptique à la membrane post-synaptique déclenchant ainsi un influx nerveux. On peut classer les synapses chimiques en deux catégories, suivant leurs fonctions: les synapses "à ruban" et les synapses "superficielles".
- Les synapses "à ruban" connectent le photorécepteur à trois éléments post-synaptiques: une dendrite (prolongement d'un neurone qui conduit le message nerveux du neurone vers d'autres cellules) de cellule bipolaire "invaginée" (BipI) au centre, et deux dendrites de cellules horizontales de chaque côté. Le pied du cône étant de grande taille, cette triade peut se répéter jusqu'à 15 ou 20 fois.
- Les synapses "superficielles", elles, ne contactent que des cellules bipolaires, mais pas n'importe lesquelles: nous venons de voir que les synapses "à ruban" ne contactent que des cellules bipolaires "invaginées" (BipI); à l'inverse, les synapses "superficielles", comme leur nom le suggère, n'impliquent que des cellules bipolaires "superficielles" (BipS).
- L'autre partie du photorécepteur est, nous l'avons déjà dit, son segment externe. Il est formé de l'empilement de plusieurs centaines de lamelles, elles-mêmes correspondant à des repliements de la membrane plasmique qui enveloppe le photorécepteur. C'est au niveau de cette partie du cône, le segment externe, que se produit l'interaction avec la lumière; les molécules impliquées, les iodopsines, sont présentes dans la membrane des lamelles.
- Le système des cônes est un système à haute résolution mais sa sensibilité est limitée. En 1967, un biologiste nommé Tornita enregistre l'activité électrique de 142 cônes de carpe; ceux ci sont éclairés par des éclairs de lumière monochromatique dont il fait varier la longueur d'onde entre 400 nm (violet) et 700 nm (rouge). Il remarque que certains cônes ont une réaction électrique maximale quand ils sont éclairés avec des radiations bleues, d'autres ont une sensibilité maximale avec des radiations vertes, et une troisième catégorie qui présente une réponse maximale aux radiations rouges. Ces résultats semblent indiquer l'existence de trois sortes de cônes. D'autres biologistes ont également pu mettre en évidence trois sortes de cônes dans la rétine humaine, ayant une absorption maximale, une dans le bleu-violet, plus exactement à 420 nanomètres, la deuxième dans le vert, à 530 nm, et la troisième à 565 nm, dans le jaune-rouge. Ces résultats seront confirmés par l'extraction de trois sortes de pigments des cônes de rétines humaines: un pigment sensible au bleu, le deuxième sensible au vert, le troisième au rouge.
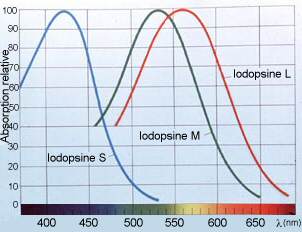
Ces spécificités ont permis de classer les cônes humains en trois catégories:
- Les cônes "S", qui contiennent en majorité le pigment sensible au bleu.
- Les cônes "M", présentant une concentration plus importante en pigments sensibles au vert.
- Et enfin les cônes "L", porteurs du pigment sensible au rouge.
(Les lettres conventionnelles S, M et L proviennent des mots anglais Short, Medium et Long wavelenght, qui correspondent respectivement aux courtes, moyennes et grandes longueurs d'onde)
La perception des couleurs
- La lumière est probablement la partie de la nature que nous rencontrons le plus souvent, pourtant nous nous demandons pas d'où elle vient exactement. Les physiciens se sont posé cette question de nombreuses fois dans l'histoire, des philosophes grecs aux disputes entre les physiciens supportant la théorie de Newton, sur la nature corpusculaire de la lumière, et ceux partisans de la théorie de Descartes, sur sa nature ondulatoire. La réponse fut finalement donnée par la physique quantique, définie par Max Planck en 1900, et travaillée aussi par Albert Einstein. Cette théorie donne place à une dualité assez surprenante, la dualité onde-corpuscule.
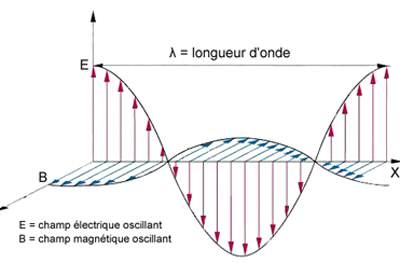
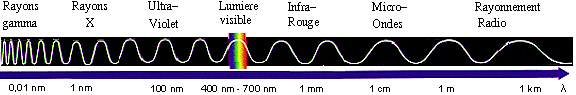
- La lumière est d'un côté une onde électromagnétique, se propageant dans l'espace et le temps. En fait, nous baignons en permanence dans un champ électromagnétique, créé par la présence de particules chargées et leur déplacement. Une perturbation de ce champ se propagera: c'est une onde électromagnétique. Ceci donne à ces ondes la propriété de pouvoir se déplacer dans le vide, car même dans le vide il y a un champ électromagnétique. Les ondes électromagnétiques sont caractérisées par leur longueur d'onde, λ, trajet parcouru par l'onde pendant une période, et leur fréquence, ν. Ceux-ci sont liés par la formule , où c est la célérité de l'onde électromagnétique, soit environ 300 000 km.s-1, que l'on appelle aussi vitesse de la lumière dans le vide. Mais la lumière visible n'est en fait qu'une fenêtre étroite de l'ensemble des ondes électromagnétiques, qui comprennent les rayons gamma, X, ultraviolets, infrarouges, les ondes radios, etc. Le spectre du visible s'étend à peu près de 400 nm (violet) à 700 nm (rouge).
- Les travaux de Max Planck et Albert Einstein sur la "lumière quantique" ont montré que l'énergie de la lumière est aussi en quelque sorte "granuleuse", ce "grain d'énergie" est appelé un photon. Chaque photon d'un rayonnement (lumière, rayons X ...) porte une quantité d'énergie caractéristique de sa fréquence. Le photon est une particule élémentaire de la famille des bosons, et explique les échanges d'énergie entre la lumière et la matière. Ces échanges avec la matière expliquent en effet comment on peut voir certains objets. Il existe deux types d'objets:
- Les objets qui produisent de la lumière, comme le Soleil, les flammes, les lampes à incandescence, etc. Ceux-ci produisent souvent de la lumière par incandescence, le mouvemement perpétuel d'agitation de la matière émettant des ondes électromagnétiques. Ces ondes ne sont pas toujours visibles, ce sont les rayonnements infrarouges, mais lorsque la température est assez élevée, elles entrent dans le spectre du visible.
- Les objets qui ne sont visibles que s'ils sont éclairés (invisibles dans l'obscurité). Ils diffusent dans toutes les directions la lumière qu'ils reçoivent, ce qui est appelé l'émission atomique ou moléculaire. En fait, un atome ou une molécule peut s'exciter par apport d'énergie de l'extérieur (par chaleur, lumière, ou décharge électrique). Dans ce cas, un électron peut "grimper" à un niveau d'énergie supérieur, mais ne tardera pas à reprendre sa place d'origine à cause de l'instabilité de cet état. Lorsqu'il reprendra sa place, il cèdera de l'énergie sous forme d'un photon émis vers l'extérieur.
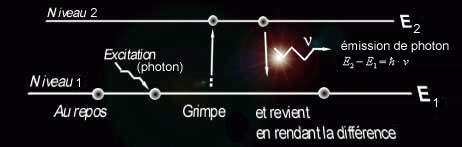
- Le photon émis portera donc une certaine énergie, qui est proportionnelle à la fréquence de la radiation, selon la formule (ħ est la constante de Planck). Selon la complexité de l'atome/de la molécule, l'émission lumineuse pourra être constituée d'un grand nombre de longueurs d'onde, matérialisées par des raies dans le spectre, ou même des bandes continues dans le cas des molécules. Une couleur est donc définie par sa longueur d'onde, ou par un mélange de longueurs d'onde. Par exemple, un vert "pur" est une radiation monochromatique de longueur d'onde 530 nm, tandis que la lumière blanche est un spectre continu contenant toutes les longueurs d'onde du domaine du visible.
- Une petite remarque sur l'infrarouge/l'ultraviolet: Pourquoi ne percevons-nous pas ces rayonnements? Deux raisons:
- Ultraviolet: tout simplement, le cristallin n'est pas assez transparent pour laisser passer ces radiations.
- Infrarouge: ici, c'est une autre affaire; l'onde parvient jusqu'au photorécepteurs, mais nous ne possédons pas de pigments visuels adaptés à cette longueur d'onde.
- Au début de la perception, nous avons donc un rayon lumineux. Celui-ci traversera les milieux transparents de l'oeil, et à cause de leurs indices de réfraction différents, sera réfracté plusieurs fois, la cornée et le cristallin assurant l'accommodation de l'image. La lentille convergente formée par ces milieux transparents provoque aussi une "inversion" de l'image sur la rétine, qui sera corrigée par l'interprétation cérébrale. Puis le rayon traverse la rétine, pour arriver finalement aux segments externes des photorécepteurs, où il sera interprété.
Gènes des couleurs
- Le rétinol des différents pigments des photorécepteurs étant le même, nous pouvons en déduire que les différences entre pigments rétiniens se font au niveau de l'opsine l'entourant. Celle-ci est une protéine, et sa séquence est donc codée par l'information génétique de l'individu.
- Ce n'est pas un gène unique qui détermine la structure des opsines: comme il en existe plusieurs différents chez un même individu, différents gènes peuvent être identifiés. Chez l'Homme, le gène codant pour la rhodopsine peut être trouvé sur le chromosome 3, celui codant pour la iodopsine S sur le chromosome 7, et les gènes des iodopsines M et L sont alignés l'un derrière l'autre sur le chromosome X, ce qu'il sera intéressant de savoir pour comprendre le daltonisme.
- La vision trichromatique n'est pas la seule possible: beaucoup d'animaux sont dichromates et certains même tétrachromates. Nous pouvons donc nous demander d'où vient le trichromatisme humain. Comme nous l'avons vu, il y a des ressemblances entre bâtonnets et iodopsine S, de même qu'entre iodopsines L et M. Selon une théorie, il y avait à l'origine deux iodopsines, S et M/L, qui se sont différenciées au cours du temps. La différenciation iodopsine S/rhodopsine est probablement plus ancienne (60% de différence) que celle des iodopsines L/M (15 acides aminés uniquement diffèrent). Les cônes S ont aussi une place spécifique dans la rétine, n'étant pas présente dans la fovéa, mais l'entourant, ce qui montre encore une fois une nette différence.
- Le trichromatisme est donc le résultat de l'existence de protéines différentes, contrôlées par des gènes.
Le daltonisme
- Forme la plus fréquente de la dyschromatopsie (trouble de la vision des couleurs), le daltonisme est une anomalie de la vue qui se caractérise par l'absence de perception des couleurs, ou par une incapacité à différencier certaines teintes ou couleurs. Ce trouble de la vision est en fait connu sous le nom de "daltonisme" depuis le 18e siècle, du nom du physicien anglais John Dalton, atteint de ce type de "cécité des couleurs" et diagnostiqué comme tel par le médecin et physicien Thomas Young. Les études menées depuis lors ont révélé les causes de cette anomalie de la vision; nous allons donc les détailler. Comme nous l'avons dit, les deux gènes qui codent pour les pigments des cônes L et M sont alignés l'un derrière l'autre sur les bras longs du chromosome X. Il en résulte, lors de la méiose (double division cellulaire aboutissant à la réduction du nombre de chromosomes, et qui se produit au moment de la formation des cellules reproductrices), des risques de recombinaisons partielles entre eux (par crossing-over : enjambement ) entraînant des perturbations de la vision colorée.
- En pratique, deux cas sont possibles: le crossing-over intergénique, et le crossing-over intragénique. Dans le premier cas, un gène entier est absent d'un des deux chromosomes; l'homme qui hérite de ce chromosome est dichromate. L'autre chromosome X, en revanche, possède le gène en double exemplaire, et l'homme qui en hérite a une vision colorée normale: le gène supplémentaire n'a pas d'effet apparent. Dans le second cas, toujours lors de la méiose, le point de rupture du chromosome se fait au milieu du gène lui même, ce qui a pour effet de séparer les exons. Le crossing-over produit alors des gènes hybrides; une partie des exons codant la iodopsine M est transférée dans le gène des iodopsines L, et vice versa. Les substitutions d'acides aminés qui en résultent modifient les courbes d'absorption des pigments de façon variée, d'où des altérations de la vision colorée qui vont d'une anomalie minime au dichromatisme.
On sait classer les formes de daltonisme; cette classification se fait selon le type du cône atteint. Aux trois types de cônes correspondent donc trois types de daltonismes:
- pathologie du cône L: le sujet est protanope (n'a pas de iodopsine L) ou protanomal (le rouge est présent mais déficient).
- cône M: la version du daltonisme la plus fréquente est la deutéranopie (manque de la iodopsine M). La deutéranomalie est la déficience du vert.
- cône S: certains sujets sont tritanopes (pas de iodopsine S), mais ce cas est extrêmement rare. Encore une fois, il existe la tritanomalie, déficience du bleu.
- Ainsi, deux daltonismes existent. D'un côté, le manque d'une iodopsine: le sujet est donc dichromate; ceci est lié au premier cas des mutations, où le gène est absent. De l'autre côté, le trichromatisme anormal, une déficience d'une couleur. Ceci est lié au cas numéro deux: le crossing-over.
- Nous avons donc vu qu'en général, le daltonisme s'explique par la présence d'un gène anormal sur le chromosome X. Chez la femme, qui possède deux chromosomes X, la présence d'un gène anormal est le plus souvent compensée par un gène normal sur le second chromosome X. La femme peut donc être porteuse du gène du daltonisme et le transmettre à ses enfants, sans pour autant être atteinte de ce trouble. Chez l'homme, qui possède un chromosome X et un chromosome Y, le gène anormal transmis génétiquement ne peut être compensé. C'est pourquoi ce trouble affecte une proportion plus élevée d'hommes: la fréquence du daltonisme est de 7 % chez les hommes et seulement de 0,5 % (7% x 7%) chez les femmes.
- Mais une dernière question subsiste: comment cela affecte-t-il la vie des daltoniens? Encore une fois, deux cas possibles. Les trichromates anormaux ont quelques erreurs dans leur vision des couleurs, mais celles-ci ne sont pas remarquables, car ils restent trichromates. Par contre, ceux qui sont dichromates (protanopes, deutéranopes et tritanopes) ont une vision très différente du monde dans lequel nous vivons. On dit souvent que les daltoniens confondent le vert et le rouge. Mais en fait, leur perception du monde coloré est très différente de celle de la plupart des gens. Leur environnement ne comporte que deux couleurs dominantes (le plus souvent bleu et jaune), mais toutes les autres couleurs du spectre leurs sont visible par des nuances différentes. Si cela est une gène pour le dichromate, c'est parce que le monde dans lequel nous vivons est adapté aux trichromates; il devient donc difficile pour les daltoniens de s'y adapter.